- El enlace peptídico.
- Restricciónes conformacionales de los polipéptidos: ángulo omega, ángulos phi y psi, ángulos chi1, chi2,…
- Hélices.
- Láminas
- Giros y bucles.

Nobel Prize in Chemistry 1954 to Linus Pauling
for his research into the nature of the chemical bond and its application to the elucidation of the structure of complex substance. Chemist, Peace Activist and something else. Linus Pauling, has won two undivided Nobel Prizes. In 1954 the Prize for Chemistry. In 1964 the Peace Prize for his opposition to weapons of mass destruction On the other hand, Pauling is largely responsible for the widespread misbelief that high doses of vitamin C are effective against colds and other illnesses.
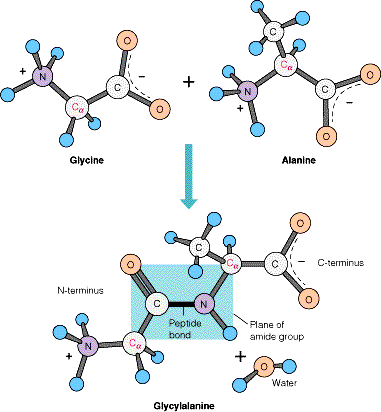
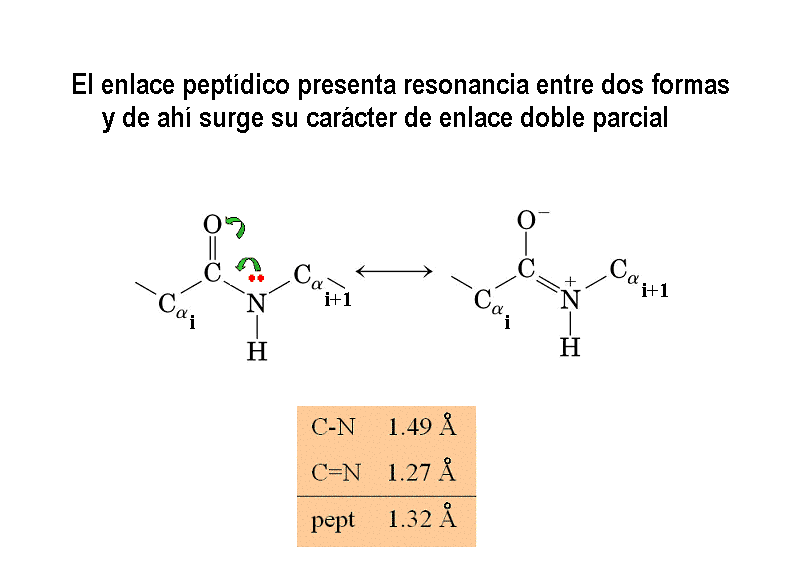
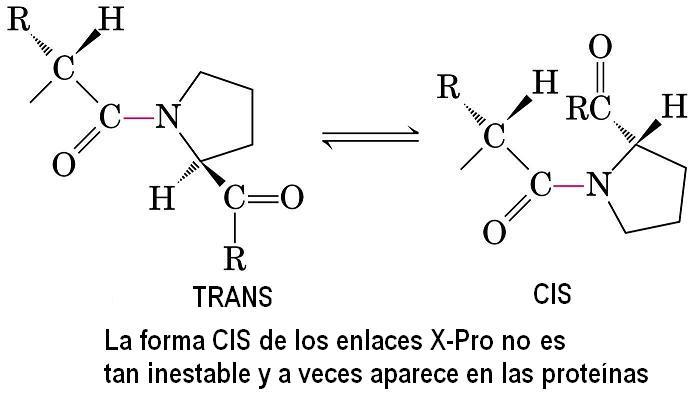
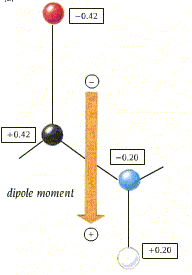
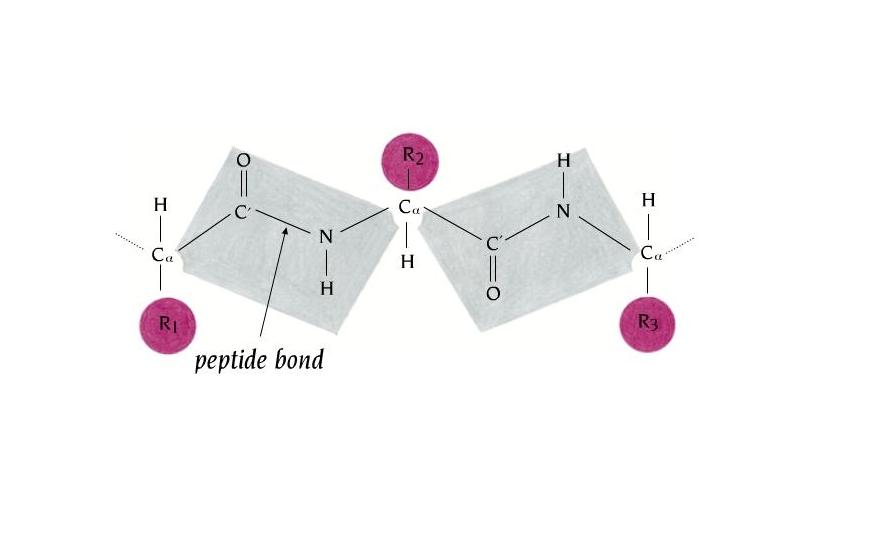
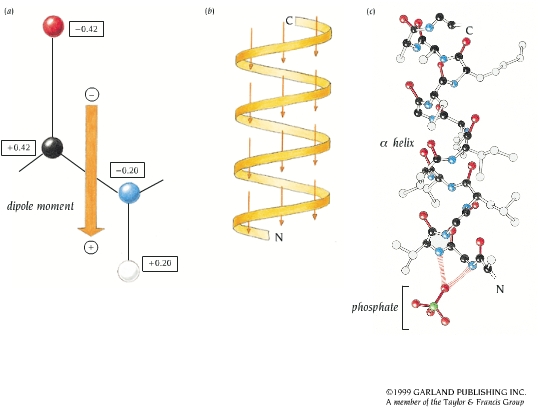
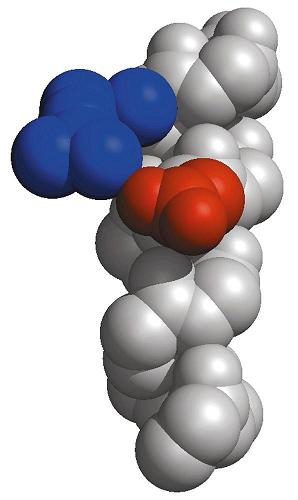
- El enlace peptídico. El enlace peptídico es el resultado de la condensación del grupo carboxilo de un aminoácido y el grupo amino de otro, con liberación de una molécula de agua. Es un enlace de tipo amida con resonancia entre dos formas. Al tener carácter de doble enlace, los sustituyentes de los átomos del enlace están en un plano. Presenta isomería cis/trans pero prácticamente siempre está en trans, salvo los enlaces X-Pro que pueden ser cis o trans. El plano peptídico tiene momento dipolar.
- Restricciónes conformacionales de los polipéptidos.
2.1. Ángulo omega: planaridad de la unidad peptídica. El giro en torno al enlace peptídico se mide mediante el ángulo omega. En enlaces cis omega vale 0, y en enlaces trans vale 180. Los demás valores no están permitidos pues se rompe la resonancia del enlace peptídico.
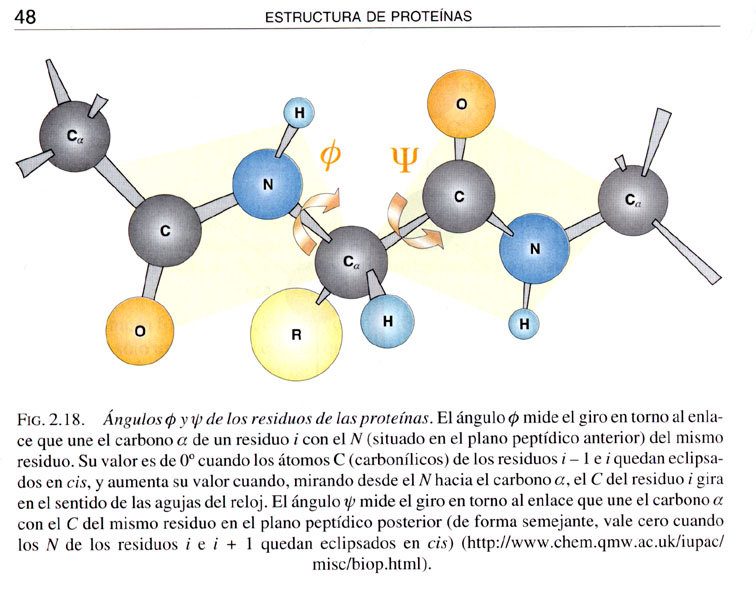
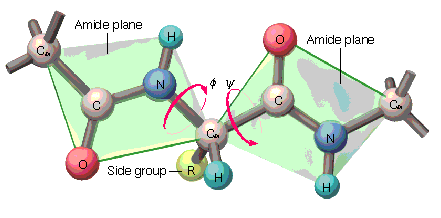
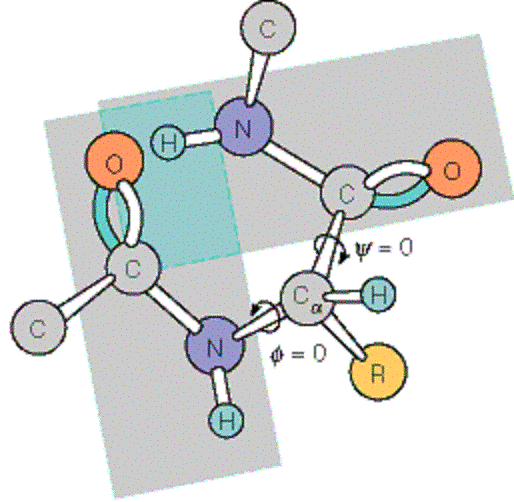
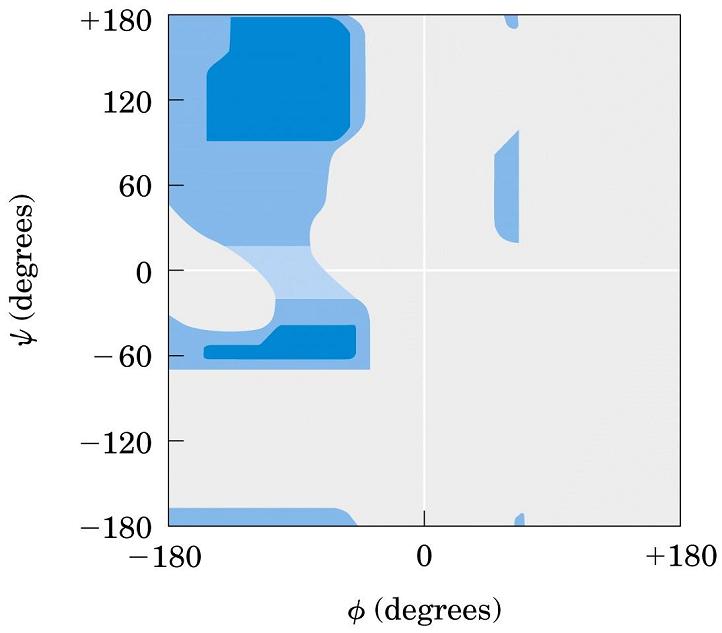
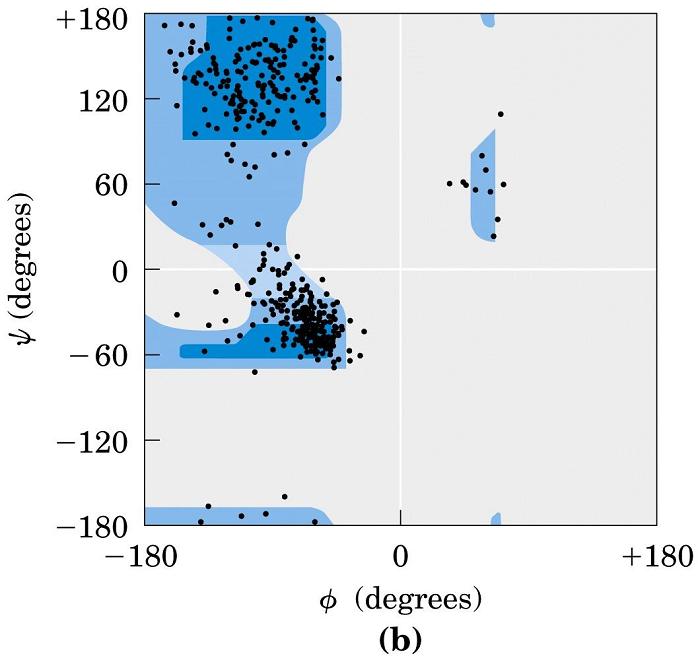
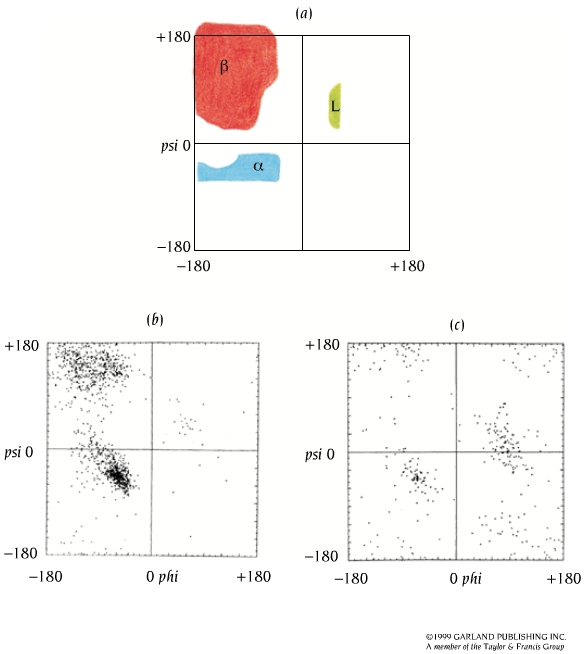
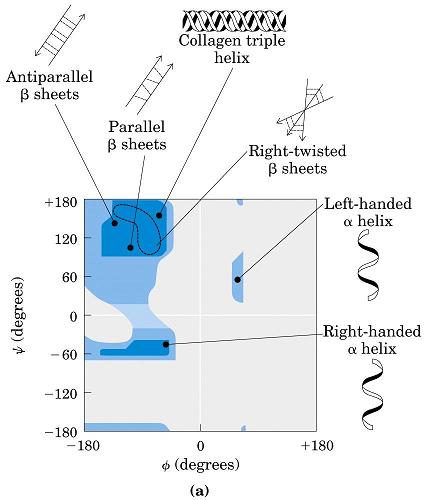
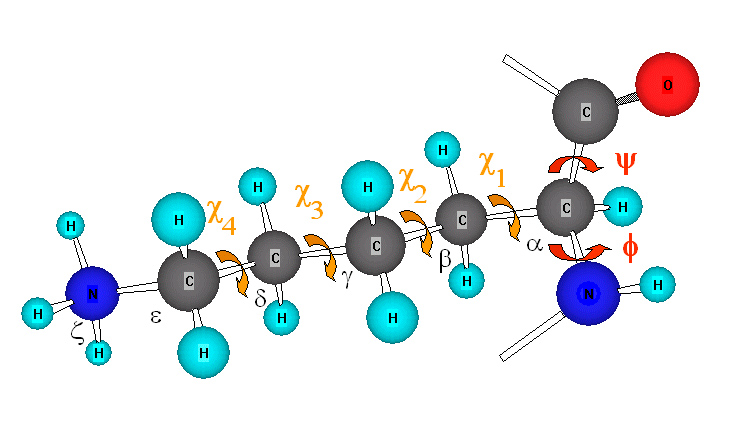
2.2. Ángulos phi y psi: diagrama de Ramachandran. Un polipéptido puede ser considerado como una sucesión de planos peptídicos. El giro en torno al enlace que une al C alfa con el N del plano anterior se mide con el ángulo phi. El giro en torno al enlace que une al C alfa con el C del plano posterior se mide con el ángulo psi. La conformación global del polipéptido queda definida por los ángulos phi y psi de cada carbono alfa. No todas las parejas de valores phi y psi están permitidas pues algunas provocarían choques estéricos. El diagrama de Ramachandran define las parejas de valores de phi y psi que están permitidas. La gran mayoría de los aminoácidos en las proteínas se sitúan en las regiones permitidas. Sólo las glicinas presentan regiones permitidas más extensas. Los elementos de estructura secundaria de las proteínas (hélices, láminas, giros) están constituidos por residuos de aminoácidos con ángulos phi y psi característicos.
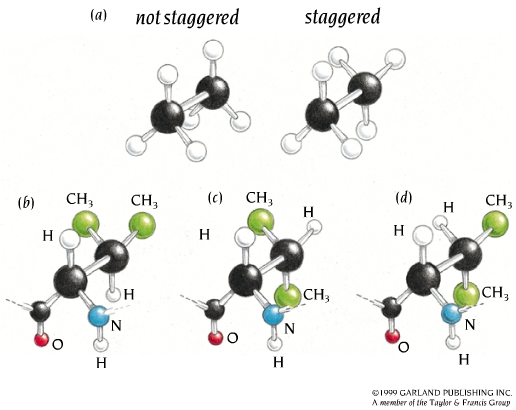
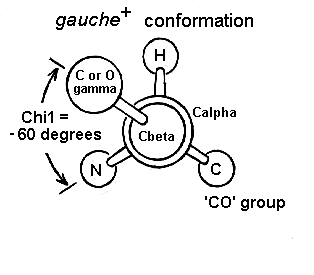
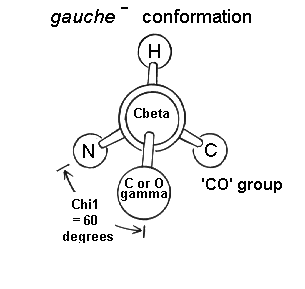
2.3 Angulos chi: rotámeros más estables. El giro en torno al enlace que une el carbono alfa con el siguiente carbono de la cadena lateral se mide con el ángulo chi1. Los sucesivos ángulos de torsión de la cadena lateral se denominan chi2, chi3,… Hay tres conformaciones alternadas del ángulo chi1 que se denominan gauche+, gauche– y trans. Domina la conformación gauche+ y despues la trans.
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
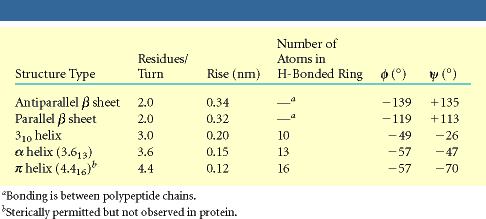
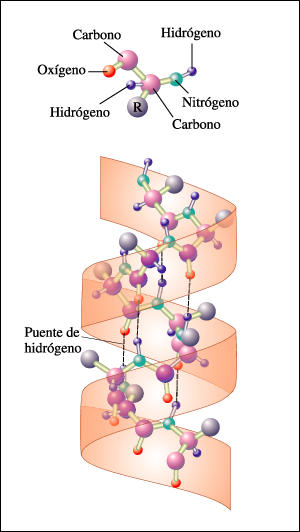
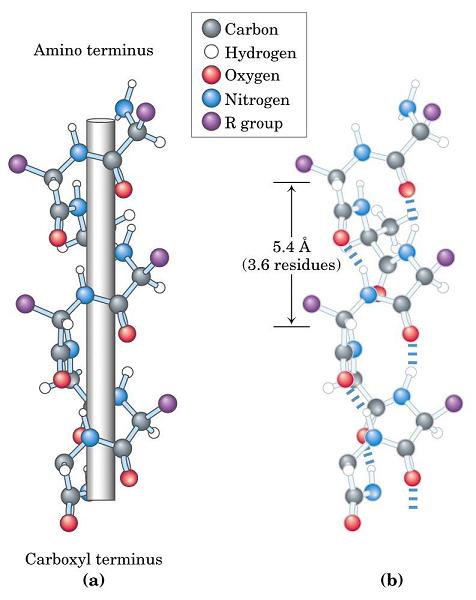
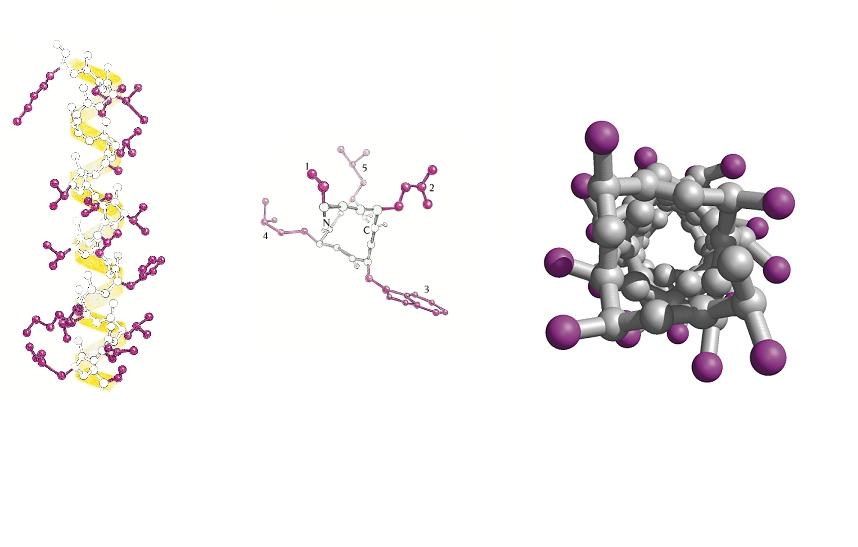
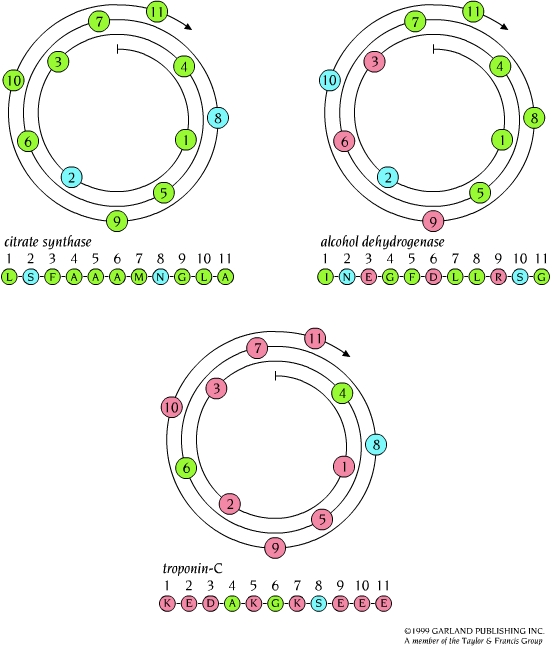
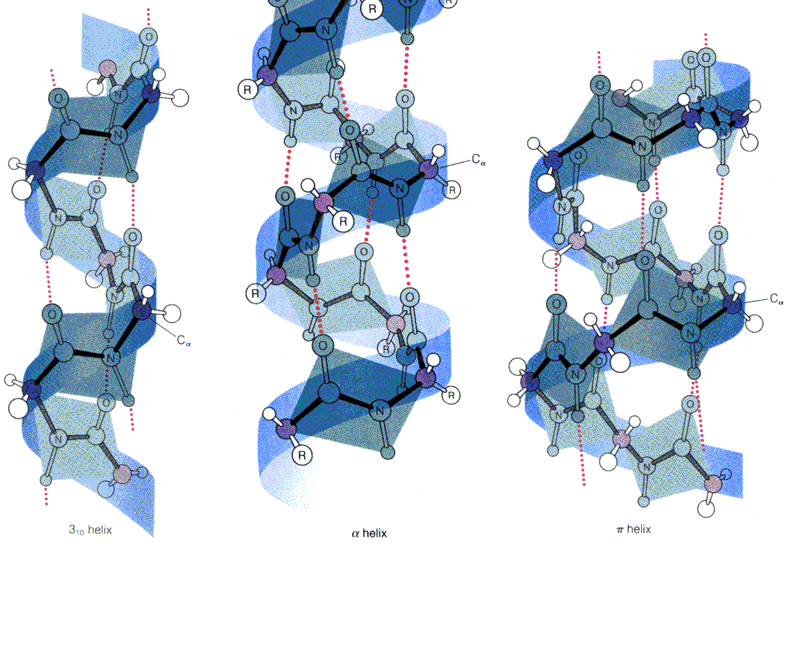
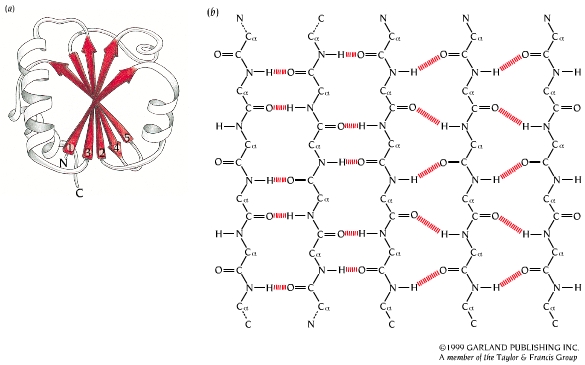
- Hélices. Cuando 5 o más aminoácidos consecutivos de una proteína adoptan ángulos phi de ~ -60 y psi de ~ -50 aparece una hélice alfa. En las hélices alfa el carbonilo (-CO) del residuo i forma un puente de hidrógeno con el NH del residuo i+4 (salvo los 4 CO del extremo C de la hélice y los 4 NH del extremo N). Las hélices alfa dan una vuelta cada 3.6 aminoácidos, avanzando 0.54 nm a lo largo de su eje (cada residuo avanza 1.5 Å). Como los planos de los enlaces peptídicos son casi paralelos al eje de la hélice, los dipolos de los enlaces peptídicos están alineados. Las cadenas laterales salen del «cilindro» de la hélice apuntando ligeramente hacia el extremo amino. Las hélices alfa de las proteínas, formadas por L-aminoácidos, son R. Las hélices (no transmembranales) tienen una longitud media de 12 aminoácidos. ¿Y las transmembranales? La estabilidad de una hélice alfa depende de: los aminoácidos que la componen, de las interacciones que se puedan establecer entre cadenas laterales a i, i+3 o a i,i+4, y de las interacciones de empaquetamiento con el resto de la proteína. Los aminoácidos que estabilizan las hélices alfa aparecen en ellas con más frecuencia que otros aminoácidos. El grado de exposición al disolvente de una hélice alfa de una proteína se puede deducir fácilmente utilizando la representación de la rueda helicoidal. Además de las hélices alfa, un polipéptido puede, en principio adoptar otras conformaciones helicoidales: las hélices con puentes de hidrógeno entre los aminoácidos i e i+3 se denominan hélices 310 (aparecen en ocasiones en el extremo C de las hélices alfa); Las hélices con puentes de hidrógeno entre los aminoácidos i e i+ 5 se denominan hélices pi (son hipotéticas).
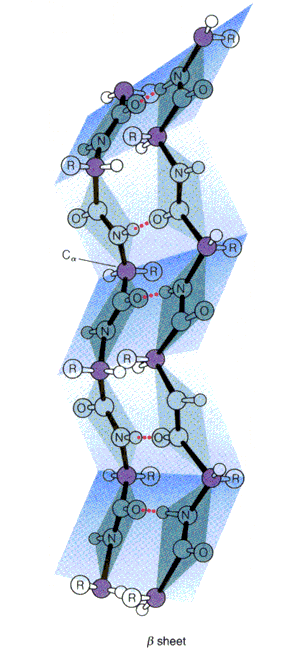
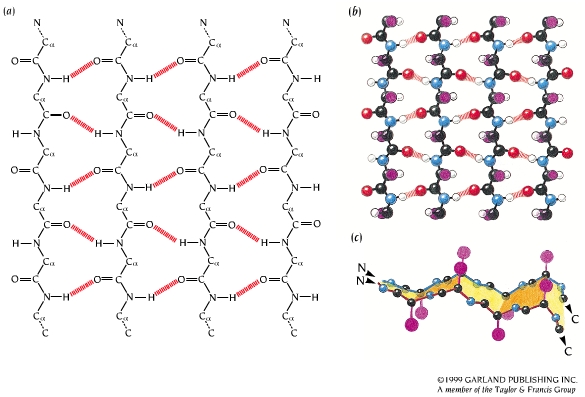
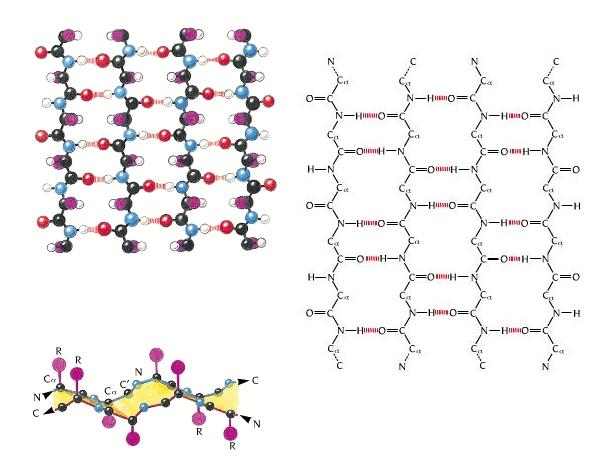
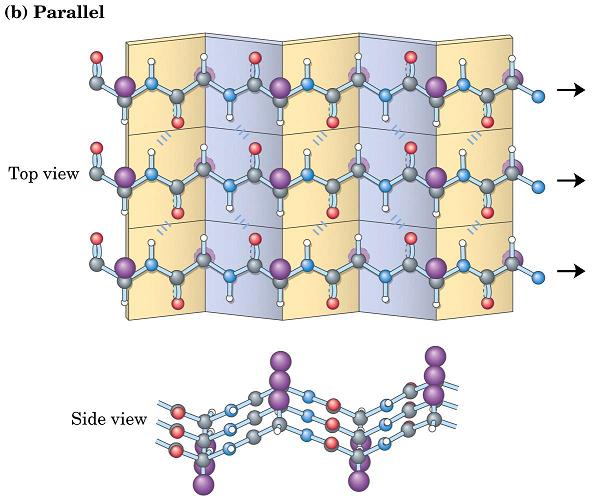
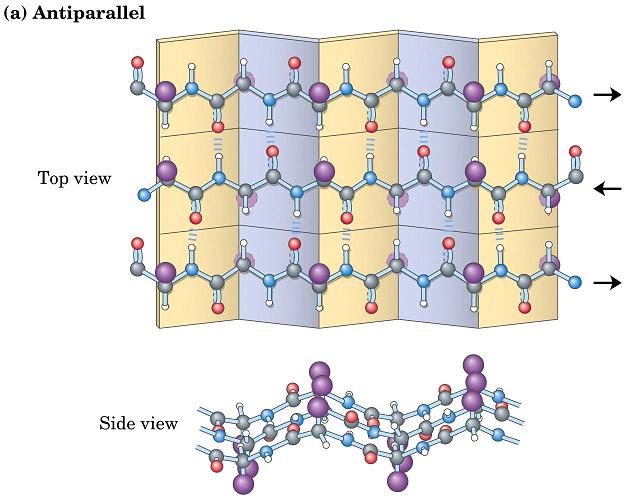
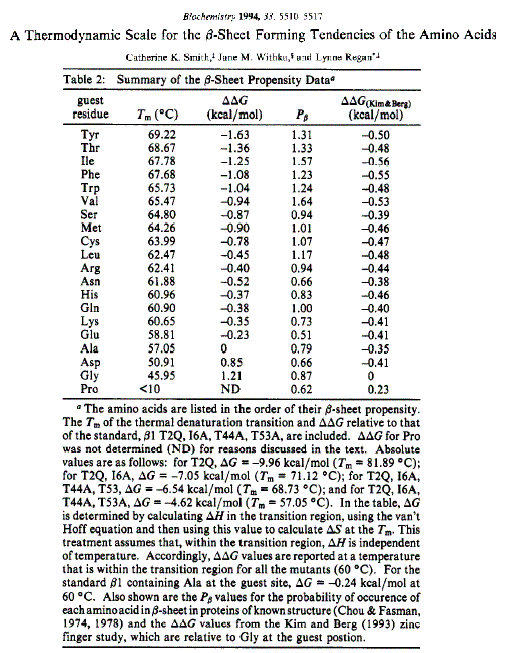
- Láminas. Cuando 2 o más aminoácidos consecutivos de una proteína adoptan ángulos phi de ~ -140 y psi de ~ +130, aparece una hebra beta (cada residuo avanza 3.5 Å a lo largo del eje de la hebra). Cuando 2 o más hebras beta se sitúan una al lado de la otra y establecen p de H entre ellas, aparece una lámina beta. Todos los CO y NH forman p de H con los de hebras adjacentes, salvo los que definen los bordes izdo y dcho de la lámina. Las láminas beta pueden ser paralelas, antiparalelas o mixtas. Las paralelas y las antiparalelas son las más frecuentes. Las hebras suelen formar ángulos de 25 ° por lo que las láminas aparecen alabeadas a izquierdas. El alabeo es mayor en las antiparalelas. Las hebras tienen un promedio de 5 residuos. Las láminas tienen un promedio de 5 hebras. Las láminas beta se estabilizan en función de: los aminoácidos que las componen, de las interacciones que se puedan establecer entre cadenas laterales de aminoácidos en hebras adyacentes, y del empaquetamiento con el resto de la proteína.
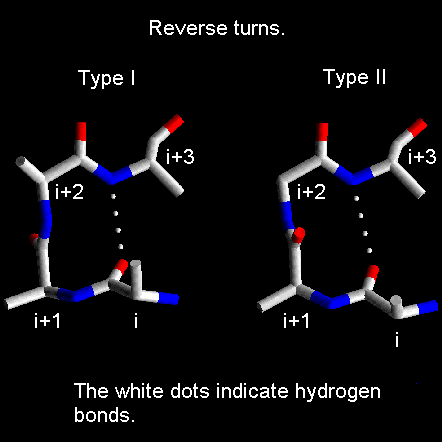
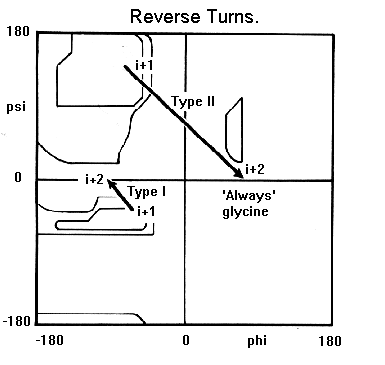
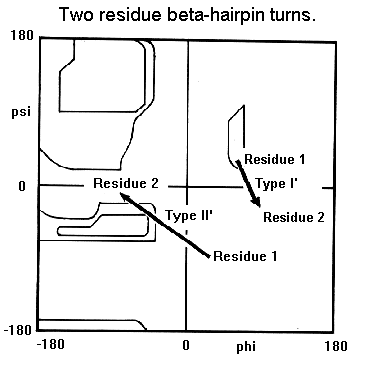
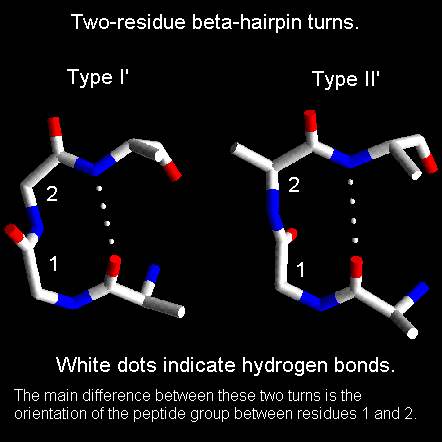
- Giros y bucles. Los giros reversos son regiones del polipéptido con un p de H entre el CO (i) y el NH (i+3) (se excluyen las hélices 310). Hay dos tipos principales (I y II) que se diferencian en los ángulos phi y psi de los residuos i+1 e i+2. Los giros que conectan hebras de una lámina beta antiparalela tienen ángulos distintos y se dominan giros beta de tipo I’ y II’. En los giros abundan las Gly. Dos hebras beta unidas por un giro beta constituyen una horquilla beta. Los bucles son las regiones de la proteína sin un valor repetido de los ángulos phi y psi. Contienen un número variable de aminoácidos. Los giros reversos y beta son bucles de 2 aminoácidos.
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}