- La doble hélice: B-DNA
- Otras formas de doble hélice: A-DNA y Z-DNA
- Estructuras infrecuentes del DNA
- El RNA de transferencia
- Estabilidad de la doble hélice
- Desnaturalización térmica de la doble hélice

The Nobel Prize in Physiology or Medicine 1962 to Francis HC Crick, James D Watson and Maurice HF Wilkins
"For their discoveries concerning the molecular structure of nucleic acids and its significance for information transfer in living material"
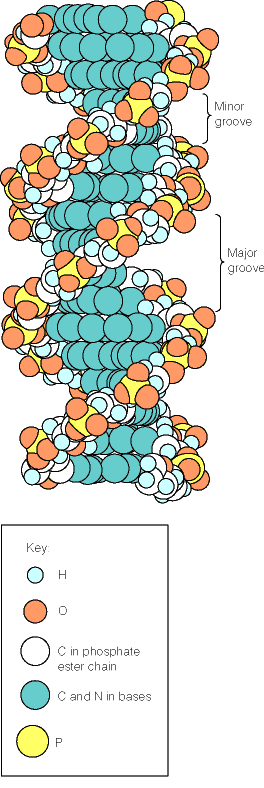
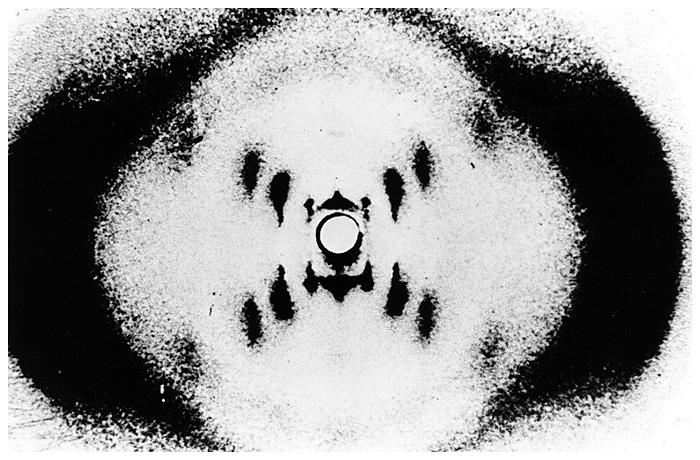
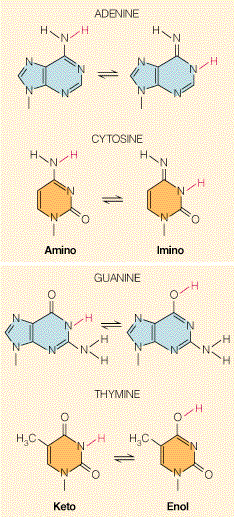
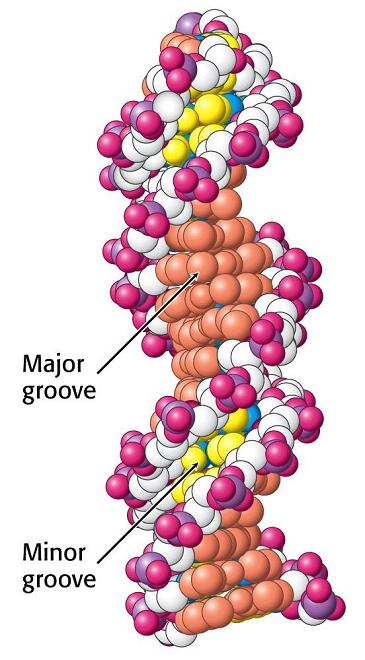
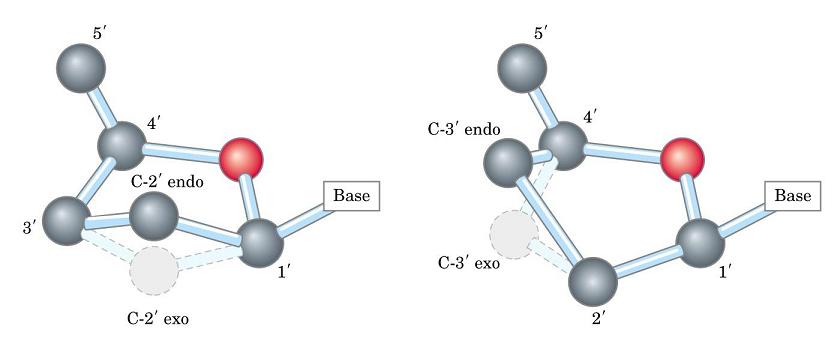
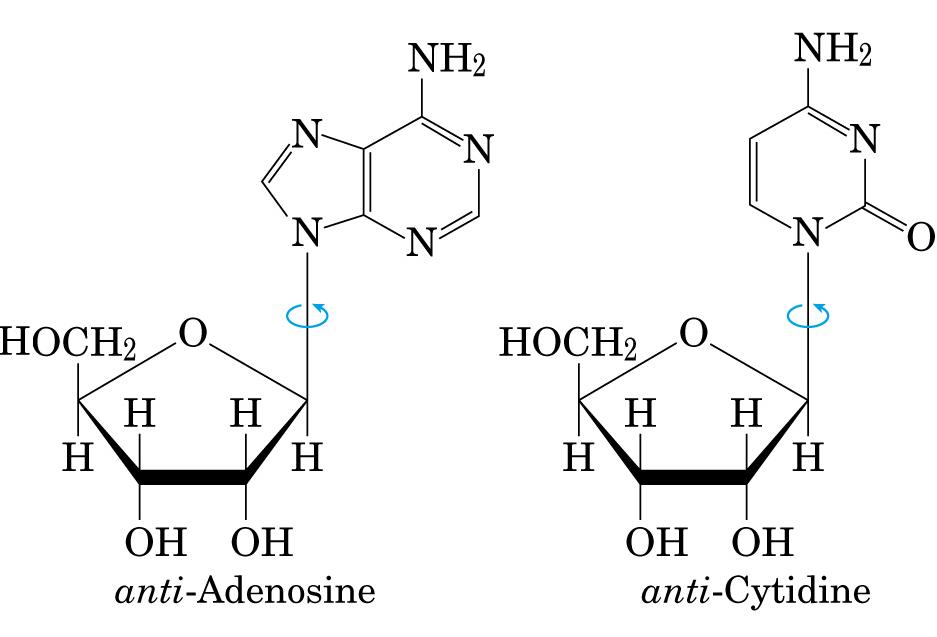
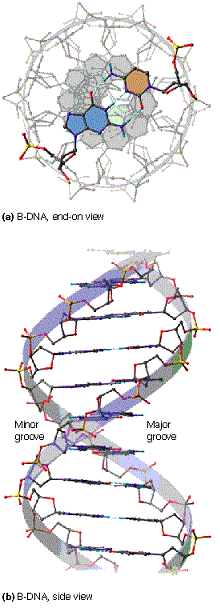
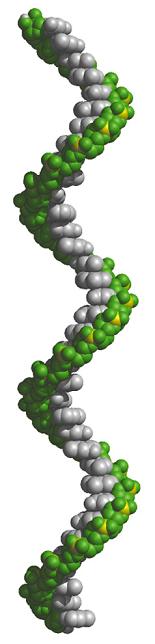
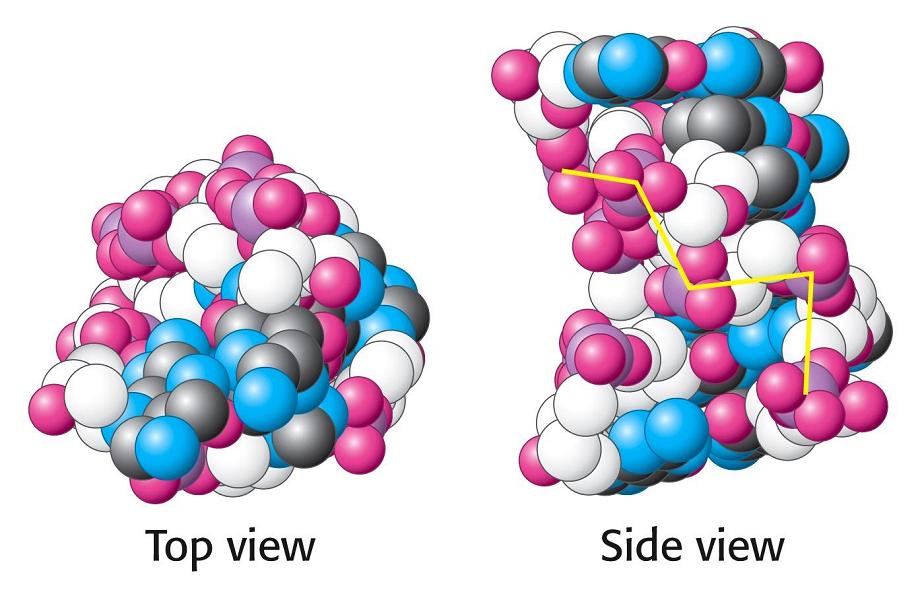
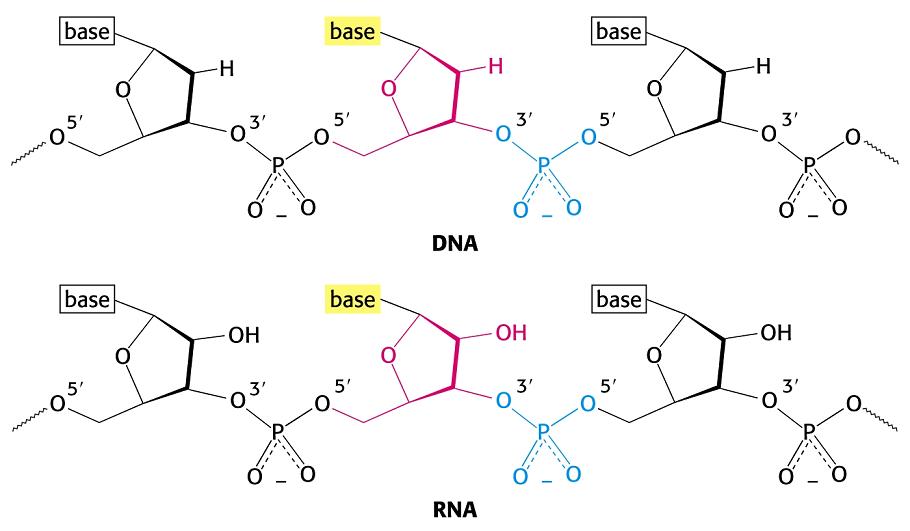
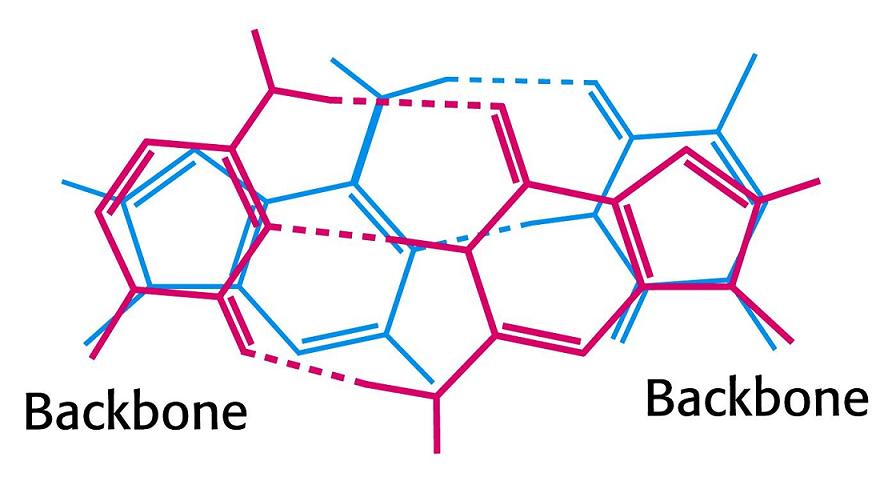
- La doble hélice: B-DNA. Es el tipo de doble hélice que predomina en el DNA. Su estructura fue modelada por Watson y Crick utilizando información de rayos x, de abundancia relativa de bases y de las estructuras tautomericas de las mismas. Presenta un surco mayor (donde las bases están más expuestas y pueden ser reconocidas) y otro menor. La desoxirribosa está en 2’ endo y así los fosfatos en 3’ y 5’ están máximamente alejados (a 7 Å). Las bases están en anti y son perpendiculares al eje de la hélice. No cabe agua en el interior de la doble hélice B. Datos geométricos de los tipos de hélices.
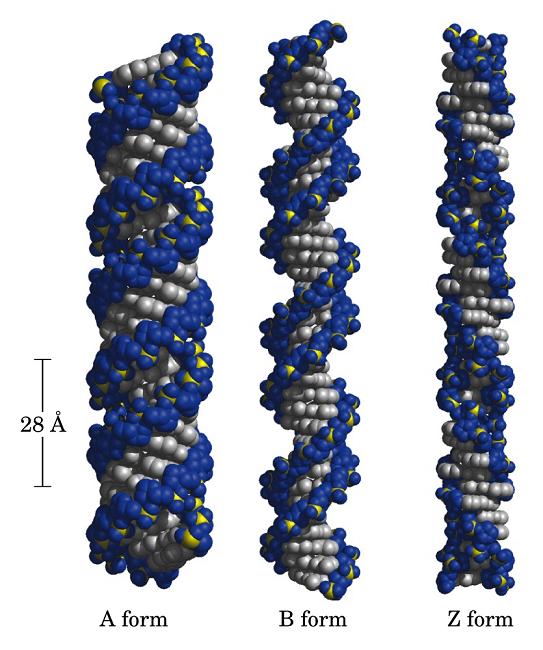
- Otras formas de doble hélice: A-DNA y Z-DNA
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
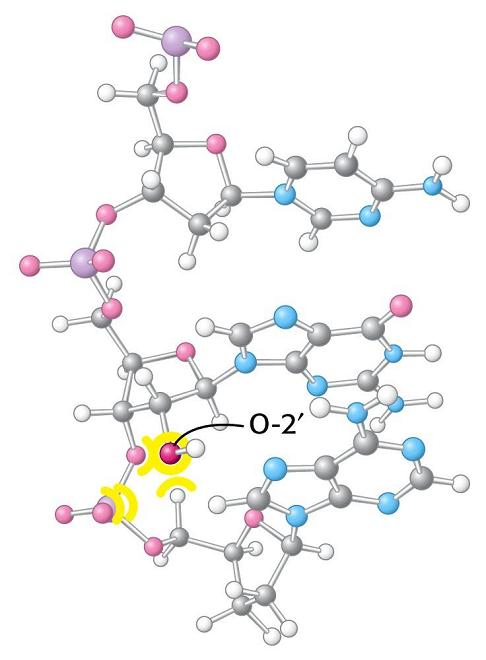
- A-DNA. En el DNA, se forma a partir de la forma B por deshidratación. Es más gruesa y corta de modo que el surco mayor es más profundo y menos accesible y en el interior de la doble hélice cabe agua. La pentosa está en 3’ endo (6 Å entre los fosfatos). Las bases no son perpendiculares al eje de la hélice. Aunque el RNA es habitualmente monocatenario, los segmentos de secuencia complementarios del RNA se aparean formando horquillas con estructura de hélice de tipo A debido a que el OH en 2’ impide la formación de la forma B.
- Z-DNA. Es levógira y, en ella, los fosfatos se sitúan en zig-zag. La forman secuencias que alternan purinas y pirimidinas (ej: poli d (GC) con poli d (CG)). Tiene un solo surco y se desconoce si aparece in vivo.
{kind=link}
{kind=link}
{kind=link}
{kind=link}
- Estructuras infrecuentes del DNA
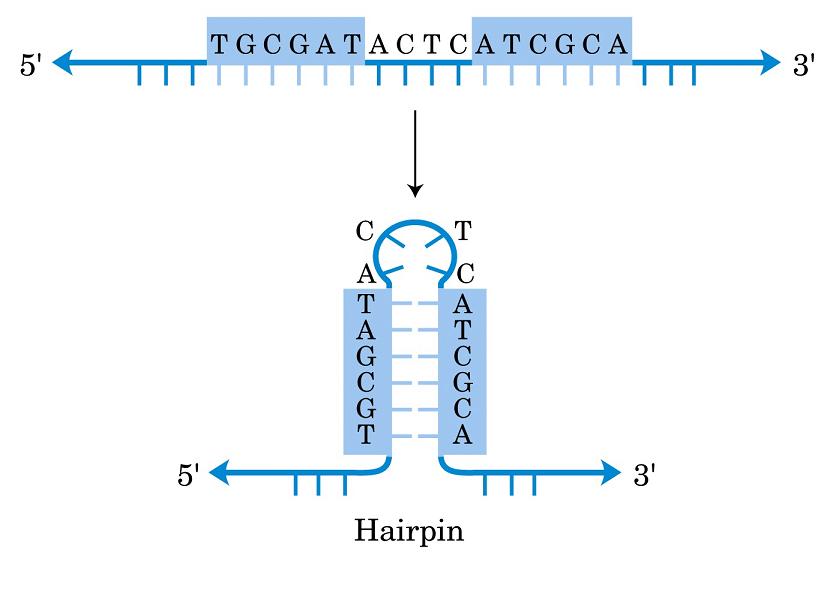
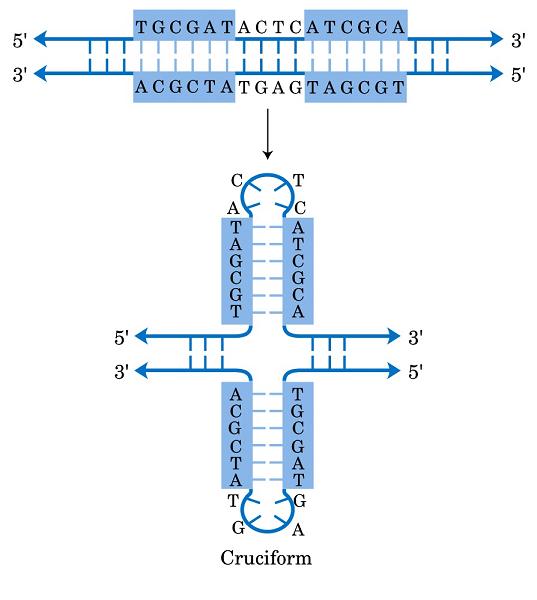
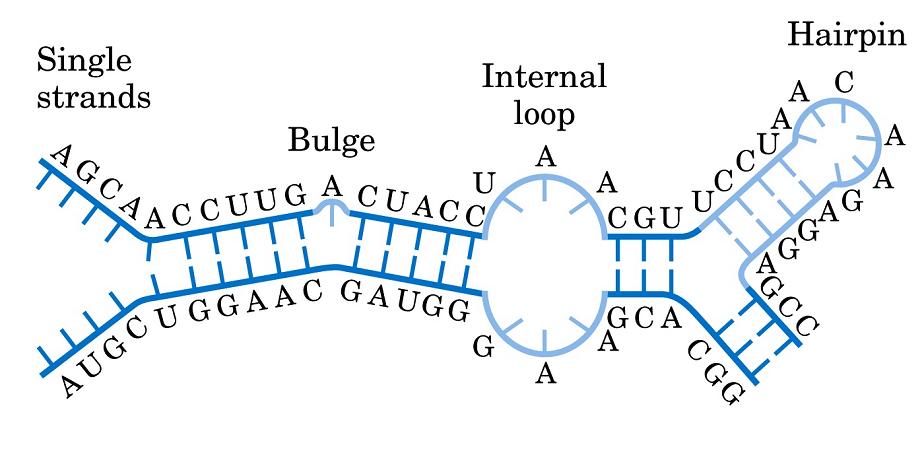
- Horquillas y estructuras cruciformes. Las estructuras cruciformes aparecen en secuencias palindrómicas e intervienen en la replicación y en la regulación de la expresión de genes. Al provocar inestabilidad en la doble hélice también se ha propuesto que podrían estar implicadas en enfermedades como cáncer o síndrome de Werner.
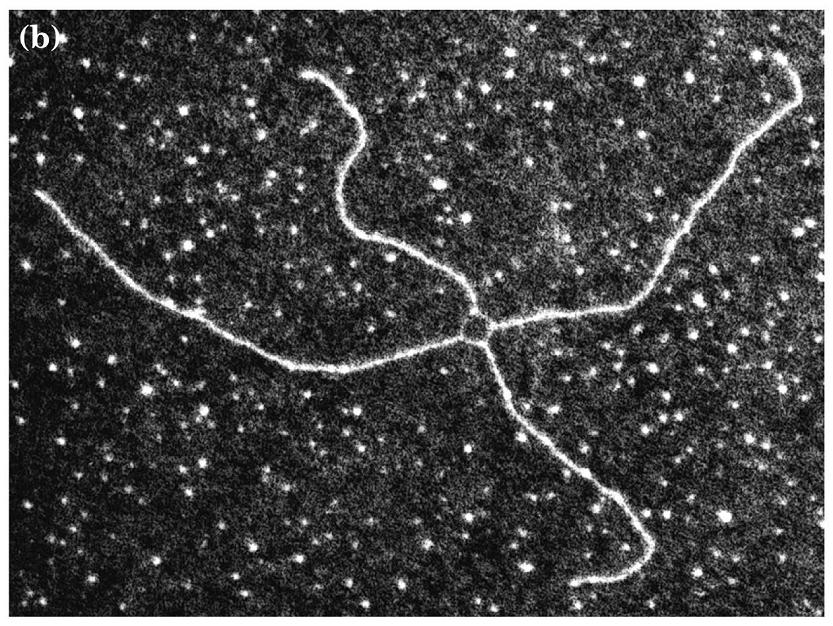
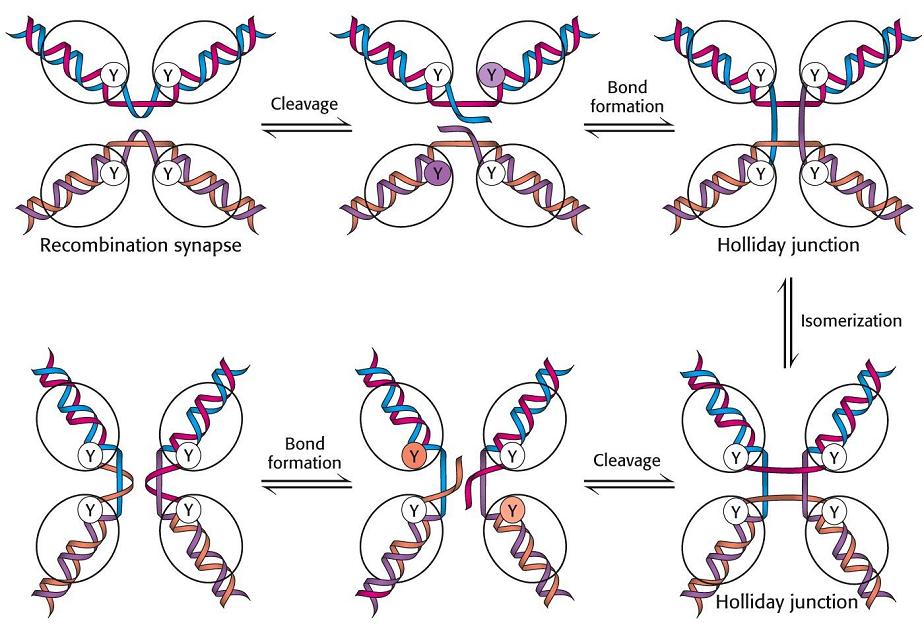
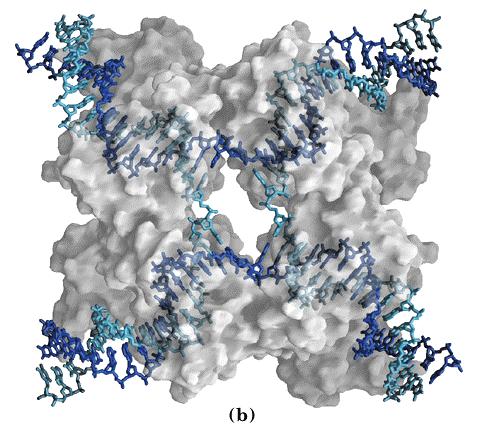
- Intersecciones Holliday. Son intermediarios de la recombinación genética.
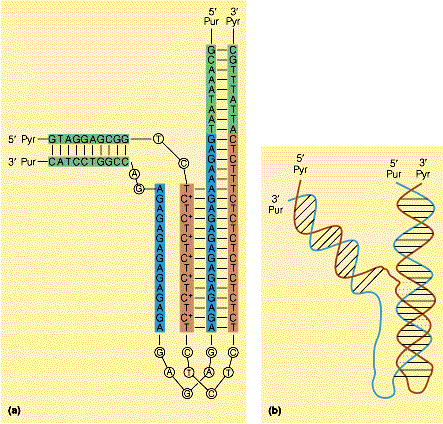
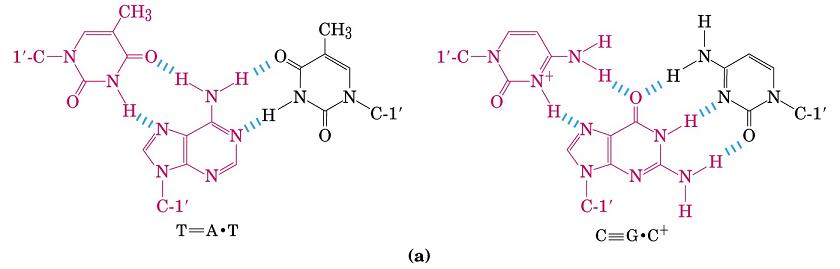
- Triples hélices (H-DNA). Aparece cuando una hebra sólo tiene purinas y la otra sólo pirimidinas. Se forman apareamientos no convencionales. El H-DNA induce represión transcripcional. Se considera una estructura explotable farmacológicamente utilizando oligonucleótidos que se unan a una determinada región y formen H-DNA.
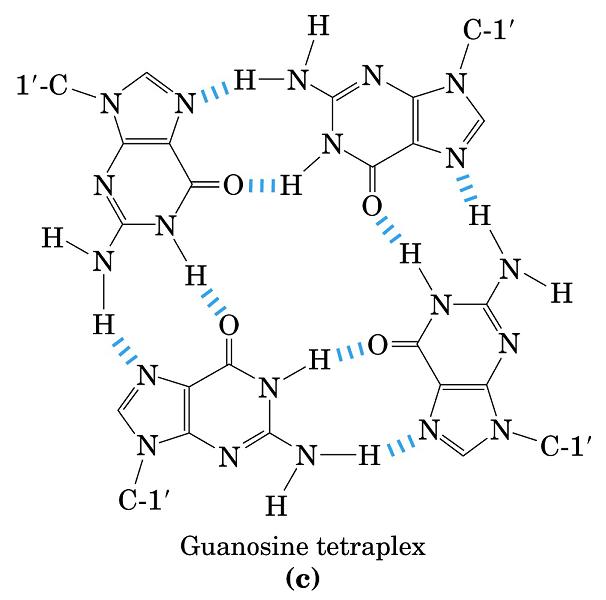
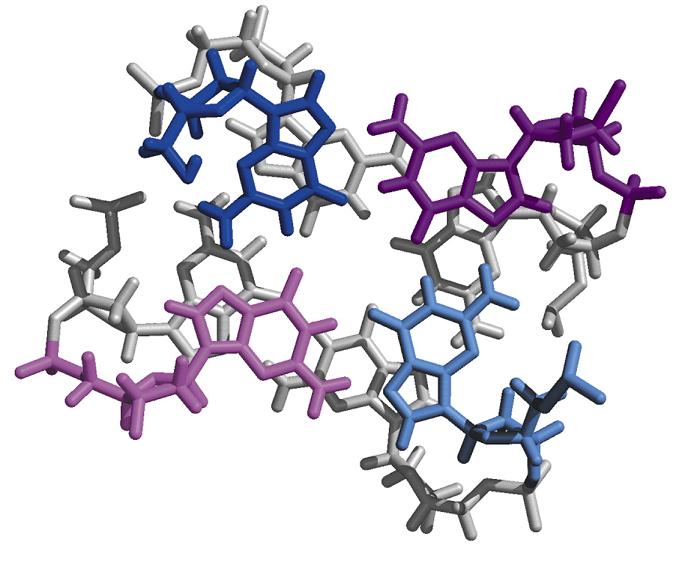
- Cadena G de los telómeros. Los telómeros aparecen en los extremos de los cromosomas eucariotas y contienen repeticiones de secuencias ricas en G (la cadena complementaria es rica en C). La cadena rica en G puede formar una hélice cuádruple. Esta estructura podría evitar la degradación del cromosoma.
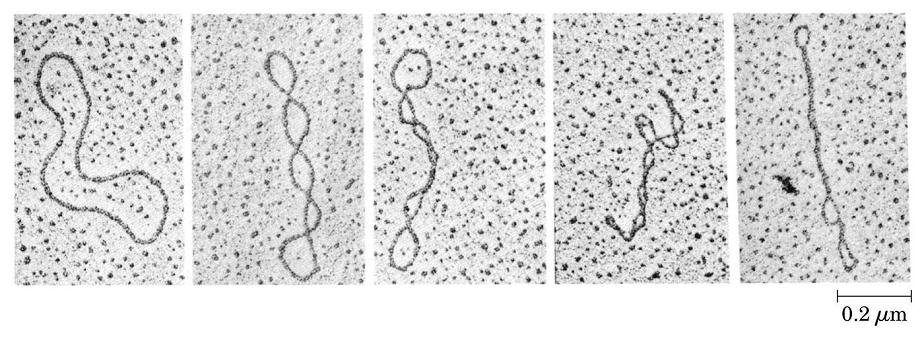
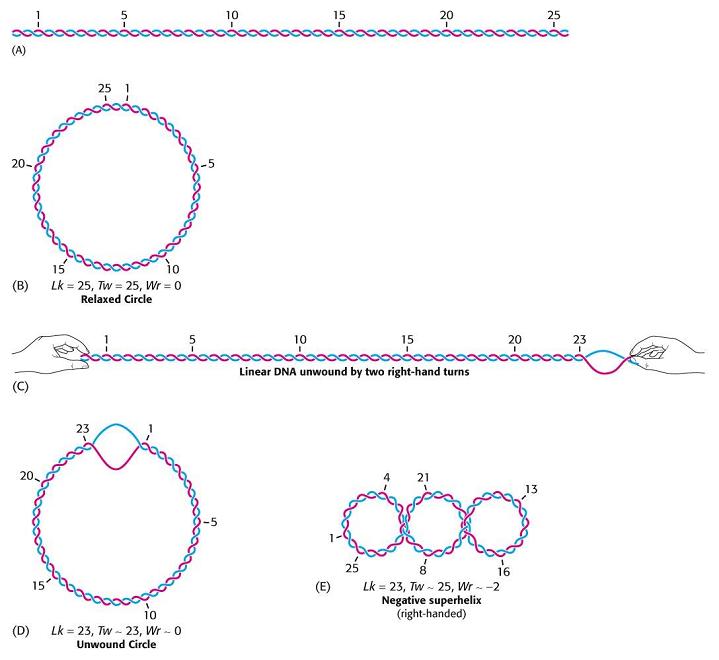
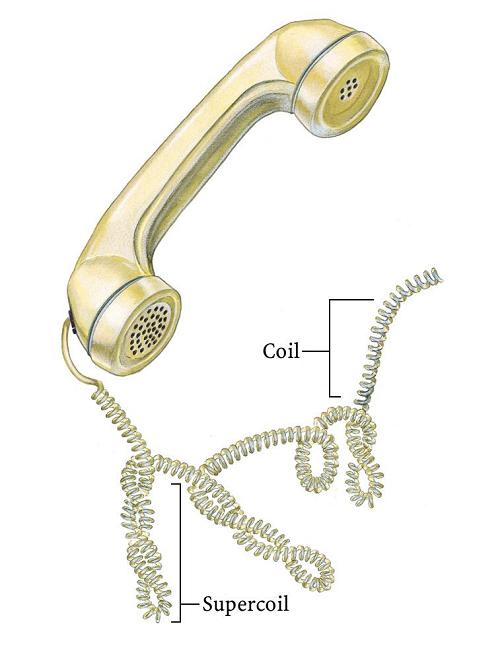
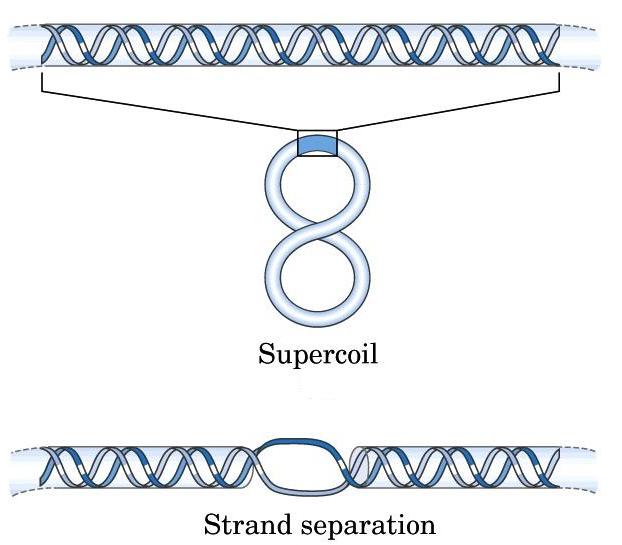
- Superenrollamiento. El DNA in vivo adopta estructuras cerradas: circulares (en bacterias) o bucles unidos por los extremos (en eucariotas). En una estructura cerrada aparece superenrollamiento cuando los extremos se unen tras haberse enroscado la cadena sobre si Como la doble hélice ya consiste en un enrollamiento de dos cadenas, el superenrollamiento de la doble hélice puede ser: negativo: en sentido opuesto al de las cadenas, aflojándolas; positivo: en el mismo sentido, apretándolas. In vivo se da fundamentalmente el negativo. El superenrollamiento negativo desestabiliza la doble hélice en 9 kcal mol-1 vuelta-1 y puede convertirse en separación de las dos cadenas, lo que podría ayudar a la replicación o a la transcripción.
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
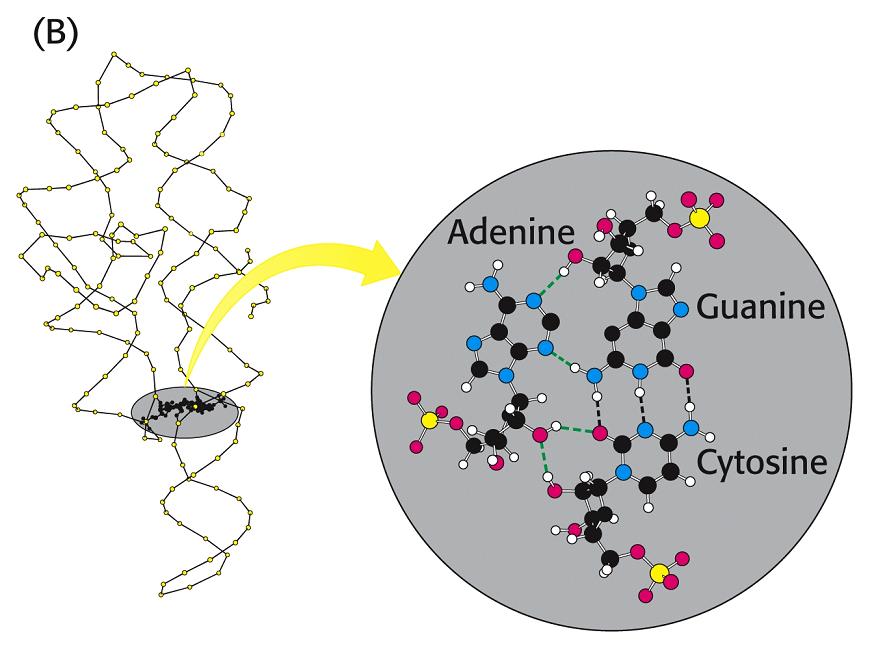
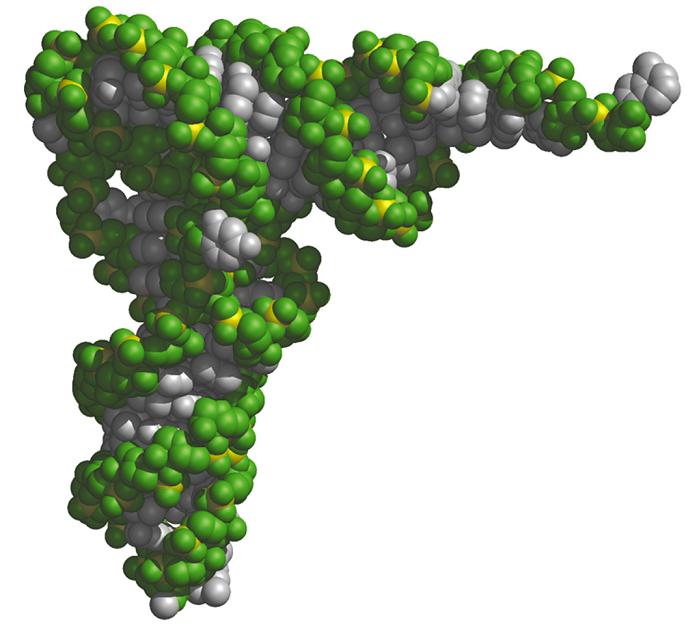
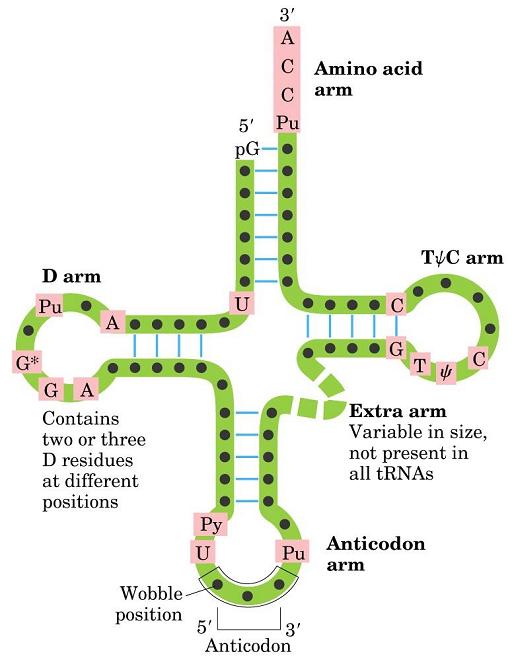
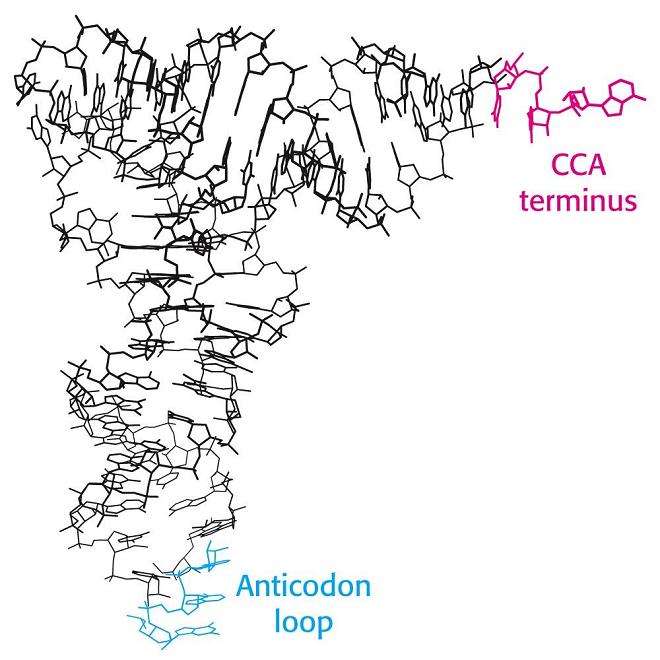
- El RNA de transferencia. En las hélices de RNA hay bucles con bases sin aparear. Además, la ribosa posee un OH en 2’ del que carece el DNA. Al poseer grupos formadores de puentes de hidrógeno sin aparear y bases hidrofóbicas sin empaquetar, algunas moleculas de RNA se pliegan y adoptan conformaciones compactas en las que esos grupos encuentran pareja. En su conformación plegada, estas moléculas de RNA presentan superficies complejas semejantes a las de las proteínas. Ejemplos de moléculas de RNA compactas son los RNAs de transferencia que deben interactuar específicamente con las aminoacil-t-RNA sintetasas, con el m-RNA y con los ribosomas. La estructura secundaria del t-RNA se asemeja a una hoja de trebol pero la tridimensional es como una «L» a uno de cuyos extremos se localiza el anticodón y al otro el aminoácido transportado.
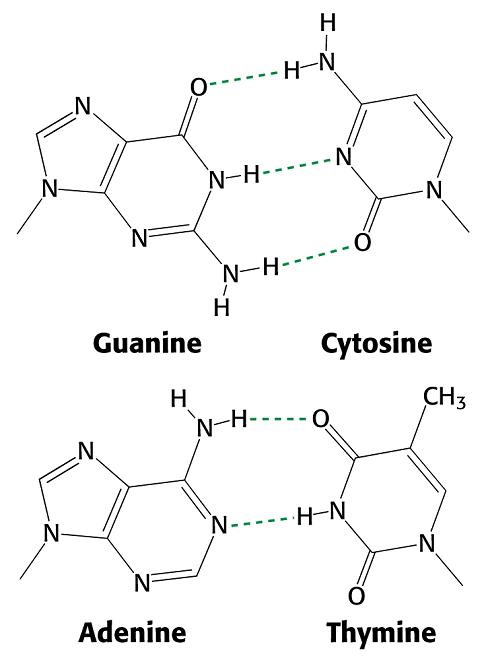
- Estabilidad de las dobles hélices. La estabilizan los puentes de hidrógeno (G—C y A–T) y el empaquetamiento entre las bases (efecto hidrofóbico, van der Waals, interaccion aromático/aromático descentrada). El empaquetamiento más favorable es entre GC/CG y el menos favorable: AA/TT. La desestabilizan las repulsiones electrostáticas entre fosfatos (contrarrestadas por unión de Mg2+ y de proteínas) y, en hélices monocatenarias (RNA), las regiones no apareadas. La estabilidad local de una molécula de ADN depende bastante del contenido AT/GC. El apareamiento AT es más fácil de distorsionar y las secuencias ricas en AT sirven a veces de sitios de unión de proteínas reguladoras.
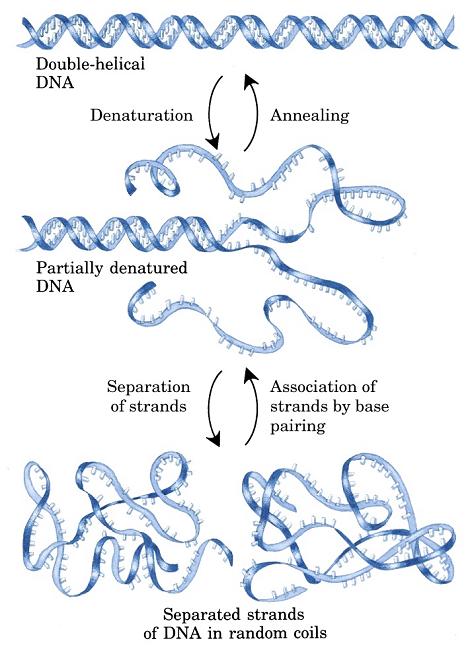
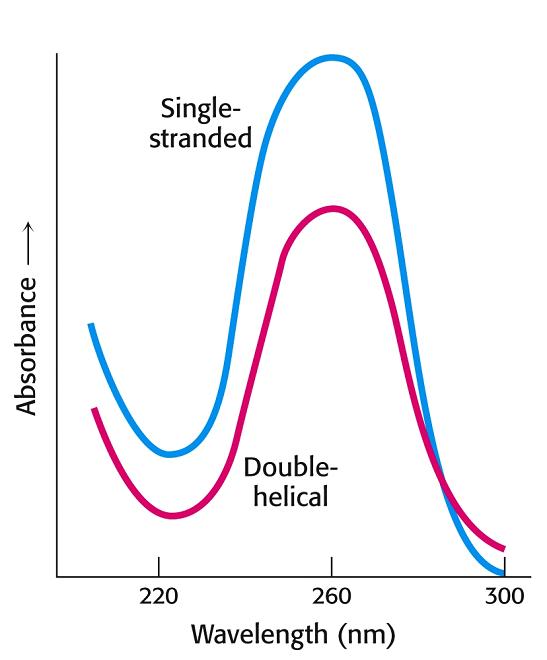
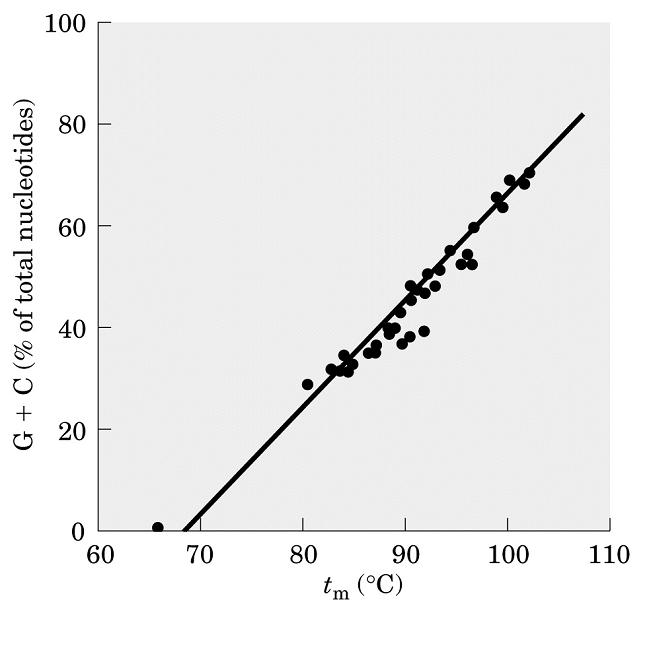
- Desnaturalización térmica de la doble hélice. Al calentar, la doble hélice se desnaturaliza y las dos cadenas se separan. Como las bases tienen mayor coeficiente de extinción en el DNA monocatenario, la absorbancia en el uv aumenta. Midiendo la absorbancia en función de la temperatura se puede calcular la Tm. La Tm del DNA es función lineal del % de bases GyC.
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}